ABSTRACT
By the methods of quantitative cytophotometry, we have identified the changes in the nucleus and of some intranuclear
compartments in the early stages of infection with encephalomyocarditis virus (EMCV). They can be characterized as
early 1 - 2 hours post infection (hpi) and temporary increase (duration about 1 hour) in the content of the acidic proteins
of the nucleolus, changing their decline to the control values. Then (after 1 - 2 hours) follows an increase in RNA content
of nucleoli to 4 hours post infection (the process takes about 2 hours). The increase in RNA content in nucleoli in
approximately the same time (slightly behind) with the activation of PML bodies (2 - 4 hpi). Then, the RNA content in
nucleoli decreased to the control values, while simultaneously decreasing activity of PML bodies (ranging from 5 - 6
hpi). The early stages of infection EMCV is also characterized by the tendency to increase in the size of the nuclei of
infected cells, and preserves at a later time. Then there is an increase in RNA content in the nucleus, roughly coinciding
with the increased content of RNA in the nucleoli.
Keywords: EMCV; Acidic Proteins; RNA; Nucleolus; Nucleus
1. Introduction
The early stages of the replication of picornaviruses represent
an extreme interest, this is due to a relatively
short period of time that the viruses which they will start
their own replication, but also establish control over a
number of important cellular metabolic processes. The
result of this phenomenon is a radical restructuring of the
entire physiology of the infected cell. Part of the mechanism
of this transformation has been clear for a long time,
while others have only recently been clarified, but much
of it is still unknown.
It is well known that picornaviruses are able to block
cap-dependent translation. Since picornaviral translation
is cap-independent by virtue of the 5 IRES, many of
these viruses have evolved potent mechanisms to inhibit
cellular cap-dependent translation during infection,
thereby thwarting detrimental antiviral responses. The
enteroviruses and aphthoviruses, for example, encode
secondary proteases at their 2A and L positions respectively,
which target eIF4G [1]. Cardioviruses do not
have secondary proteases. Their L and 2A proteins have
essential host shut-off roles, but use non-proteolytic
mechanisms to achieve them. The EMCV L (67 aa) contributes
to the inhibition of cap-dependent translation by
triggering disruption of nucleocytoplasmic trafficking
during infection. For the disruption of cap-dependent
translation of the host cells with EMCV, 2A protein is
responsible. As shown by [1], 2A NLS sequence is required
for virus shutoff of cap-dependent host protein
synthesis.
It has been shown that proteins of EMCV-the 2A, 3B
(VPg), 3C (pro) and 3D (pol) can also be found within
the nucleoli. The localization of these proteins occurs in
the first 2 - 4 hours following infection of cells [2,3].
Therefore, it is important to study the influence of the
picornaviruses on the nuclear structure of infected cells.
As it is well known, the replication of picornaviruses
occurs within the cytoplasm. However, at the earliest
stages of viral infection many viral proteins are observed
in the nucleus of affected cells. The research reported in
this paper was directed towards determining the DNA,
RNA and acidic proteins (non histone) as well as PML
bodies in nuclei of EMCV infected cells. *
Corresponding author.
Copyright © 2013 SciRes. CellBio
126 Z. A. KARALYAN ET AL.
2. Materials and Methods
2.1. Cells
SK-N-MC human neuroblastoma cells were cultured in
Dulbecco’s Modified Eagle Medium (DMEM) (Sigma)
supplemented with 10% heat-inactivated fetal bovine
serum (FBS) at 37˚C in 5% CO2.
2.2. Virus
EMCV (Columbia-SK strain) was used at multiplicity of
infection 105 TCD50/ml on SK-N-MC. Viral titers were
calculated by the method of Kärber. As a control the parallel
conducted passages of noninfected cultures were
used.
2.3. Image Cytophotometry
In order to quantitative DNA analyze of the received
data, the cells preparations were fixed in 96% ethyl alcohol
for 30 minutes and painted in fresh Shiffs reactive,
by Feulgen (hydrolyze 5N HCL 60 minutes at 22˚C). The
content of DNA in a nucleus and nucleolus was defined
by computer-equipped microscope-photometer SMP 05
(OPTON). The image cytophotometry of DNA was performed
on 575 nm wave [4]. Unstimulated human lymphocytes
were used as diploid standards.
For quantification of RNA was used gallocyanin
chromalum stain. To obtain reproducible staining results
with these large sections, the method of Einarson was
adapted to quantitative [5] and image analytical requirements.
The image cytophotometry was performed on 610
nm wave. In each case controls were evaluated as 100%.
Fast green FCF staining (for acidic proteins) was used
in Deitch modification [6,7]. The image cytophotometry
was performed on 434 nm wave. In each case controls
were evaluated as 100%.
2.4. Determination of the PML
Cells grown on glass cover slips were fixed in 4% paraformaldehyde/PBS
(pH 7.5) for 5 minutes at room temperature,
permeabilized in 0.5% Triton X-100 in PBS for
5 minutes at room temperature. PML protein was visualized
with the monoclonal antibody. The determination of
the PML was performed using monoclonal antibodies
“PML PG-M3, Santa Cruz Biotechnology Inc”-catalogue
no. sc-966 FITC [8].
2.5. Statistics
All experiments were conducted in triplicate. The significance
of virus-induced changes was evaluated by
two-tailed Student’s t-test. p values < 0.05 were considered
significant. SPSS version 15.0 software package
(SPSS Inc., Chicago, IL, USA) was used for statistical
analyses.
3. Results
The lytic EMCV infection was received by virus introduction
on 48 h confluences of SK-N-MC culture. At 8
hours post infection (hpi) the virus titer reached 3.0 lg
TCD50/ml, at 12-4.0, at 24-4.5, at 24-72-6.5 (Figure 1).
The time-course of a single cycle of EMCV reproduction
in SK-N-MC cells take place about 8-12 h.
The average content of the nucleolus to a nucleus does
not change during the whole period of the experiment
with the exception of some minuscule increases of the
content of nucleolus in a nucleus (2.5-control, 2.9-at 6
hpi). The cytometry of the nucleolus values revealed the
following results: in the whole there are not definite
changes, but there may an emphasized tendency to increase
the given value up to the 6 hpi. The total area of
the nucleoli increases almost by 35%, but due to large
variations in individual performance the difference is not
reliable.
Thus, when the infection with picornaviruses occurs
the activation of the nucleolar values is detected. It has a
temporary nature―first of all increases in the synthesis
of the acidic proteins and after that an increase in the
content of the RNA and the area of the nucleoli occurs.
The quantitative indicators of DNA of the nucleus do
not differ from the control values throughout the experiment,
nor in the DNA content and neither in the distribution
of DNA in ploidy classes.
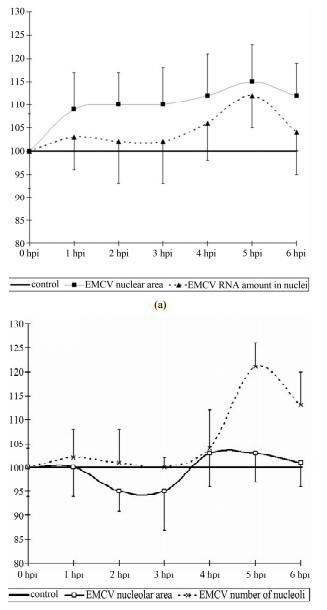
The size of the nucleus tends to increase in comparison
with the control (p < 0.1), by 1 hpi and remain so until
the end of the observation period (6 hpi) (Figure 2(a)).
In addition the increase in the size of the nucleus is small
and varies from 8% - 11% from the baseline. It is important
to note that the increase in the size of the nuclei of
infected cells is not accompanied by an increase in the
RNA or acidic proteins of the nucleus. However, if the
content of acidic proteins of the nucleus remained in a
relatively stable index throughout the study, the RNA has
a tendency (t = 1.8, p < 0.1) increased by 5hpi, and coinFigure
1. The titer of EMCV calculated on SK-B-MC cells.
Copyright © 2013 SciRes. CellBio
Z. A. KARALYAN ET AL. 127
(a)
(b)
Figure 2. Dynamics of nuclear (a) and nucleolar (b) indices
under the influence of EMCV infection. Data show percentage
of control levels (average of 300 cells = 100%).
cides with an increase in RNA in the nucleoli (Figure
2(b)).
As it follows from the Figure 3, in the early stages of
the infection, there is an increase in the content (amount,
concentration) of the nucleolar acidic proteins (with a 1-
2 hpi) (t = 3.07, p < 0.05). The elevated level of acidic
proteins has a short-period character, and after 1 hour (to
3 hpi) its levels in the nucleolus does not differ from the
benchmarks. The increased levels of the acidic proteins
vary between 20% - 25% of the initial content.
The RNA in the nucleoli of the infected and control
cells do not differ from each other, up to 4 hpi, when an
Figure 3. Dynamics of nucleolar RNA and acidic proteins
under the influence of EMCV infection. Data show percentage
of control levels (average of 300 cells = 100%).
increase in the RNA content in the nucleoli of infected
cells begins to be observed (Figure 3) (t = 2.17, p <
0.05). The increase of the RNA in the nucleolus reaches
30% - 35% in comparison with the control values. The
increase of the RNA as long with the proteins has a short
period character and finishes after 5 hpi, decreasing
down to the levels of the control values (6 hpi).
The intensity of the luminescence PML bodies, in the
nuclei of the infected cells does not differ from the background
values in the range from 1 to 2 hpi. Then there is
a sharp increase in the intensity of luminescence, which
indicates about the increased activity of PML bodies by 3
hpi. Increased activity of PML bodies completely fading
away to a 4 HPI, after which it does not differ from the
control values (Figure 4).
We have identified the changes in the nucleus and of
some intranuclear compartments in the early stages of
infection EMCV.
They can be characterized as early (1 - 2 hpi) and
temporary increase (duration about 1 hour) in the content
of the acidic proteins of the nucleolus, changing their
Copyright © 2013 SciRes. CellBio
128 Z. A. KARALYAN ET AL.
Figure 4. PML bodies in EMCV infected SK-N-MC cells. (A)
PML bodies in control nuclei of SK-N-MC cells; (B) PML
bodies in infected nuclei of SK-N-MC cells (3 h.p.i). (C)
PML bodies in infected nuclei of SK-N-MC cells (6 h.p.i.
EMCV); (D) PML bodies in infected nuclei of SK-N-MC
cells (9 h.p.i. EMCV) (×400).
decline to the control values. Then (after 1 - 2 hours)
follows an increase in RNA content of nucleoli to 4 hpi
(the process takes about 2 hours). The increase in RNA
content in nucleoli in approximately the same time
(slightly behind) with the activation of PML bodies (2 - 4
hpi). Then, the RNA content in nucleoli decreased to the
control values, while simultaneously decreasing activity
of PML bodies (ranging from 5 - 6 hpi). The early stages
of infection EMCV is also characterized by the tendency
to increase in the size of the nuclei of infected cells, and
preserves at a later time. Then there is an increase in
RNA content in the nucleus, roughly coinciding with the
increased content of RNA in the nucleoli.
The total content of acidic proteins of the nucleus is a
relatively stable index, changing a little both in the control
and in the early stages of infection EMCV.
4. Discussion
For EMCV is found the involvement of the viral proteins
in the nuclear processes. Already in the early stages of
infection (2 - 3 hpi) cardiovirus protein 2A, as well as
proteins 3BVpg, 3Cpro, 3Dpolas a single precursor
3BCD are located in the nucleoli of the cells, where the
synthesis of rRNA and ribosome assembly take place
(Aminev et al., 2003a, Aminev et al., 2003b). The function
of the cardiovirus proteins in the nucleus of infected
cells have not been studied sufficiently. Protease 3Spro
presumably responsible for the inhibition of synthesis of
cellular mRNA, whereas the work of RNA polymerases I
and III in cardiovirus infection are not inhibited [3]. The
function performed by the protein 2A of cardiovirus in
the nucleoli of the cells is not completely clear. Amines
and colleagues suggest that the protein 2A in the nucleoli
embedded in the mature ribosomal subunits, resulting in
formation of modified ribosomes engaged mainly in the
cytoplasm of virus-specific protein synthesis [2]. As
shown by us, at this stage is a series of important structural
and functional changes in the nucleoli of infected
cells.
With regard of the above said the increased content of
nucleolar acidic proteins in early stages of infection
EMCV could be explained by two reasons. The first is
the accumulation of the viral proteins and particularly
proteins 2A, 3BVpg, 3Cpro, 3Dpol and its predecessor
3BCD. The second - the accumulation of nucleolar proteins
with enzymatic activity (discussed below).We have
identified the data which allow us to assert that in the
early stages of infection (2 - 4 hpi) significant changes
occur in the nucleoli, coinciding with the localization of
viral proteins in them. These effects disappear in the later
stages of infection (6 hpi and later). This suggests a
short-term increase in the functional activity of the nucleolus
[9] under the influence of infection EMCV. In
addition to changes in the nucleoli, this process is accompanied
by an increase in the PML bodies. PML
bodies-small spherical domains are present in the nucleus
of cells, they undergo morphological changes during the
cell cycle. Their number is intensely variable, depending
on the physiological state of the cells, cell cycle stage in
viral infections and so on. The PML bodies are destroyed
during some viral infections.
Incubation of the cells with IFN induces the synthesis
of the PML-protein and inhibits the multiplication of the
viruses. These cells are destroyed during a viral infection
and for the replication of adenovirus DNA their destructtion
is a necessary step that underlines the possible involvement
of the corpuscles to provide antiviral activity
of cells. Incubation of the cells with interferon induces
the synthesis of PML-protein and inhibits proliferation of
some viruses [10]. Upon infection of cells with the virus
EMCV, it is shown a decrease in the number of the PML
bodies, under the influence of the viral 3C protease. As
the mechanism of reduction of PML bodies, the authors
consider a possible direct degradation of the PML bodies
3C protease EMCV [11]. Thus, the sharp decrease in
emission intensity of monoclonal antibodies to PML, to
5.6 HPI, is explained by the influence of non-structural
viral proteins (3Cprotease), and aimed at suppressing
IFN-mediated protection of the infected cell.
We have also investigated the acidic proteins of chromatin
and the nucleolus in a control experiment and under
the influence of the virus. Acidic proteins of the
chromatin play an important role in the regulation of the
genetic activity. It was shown that in proliferating tissues
contain more acidic proteins than that of resting, and
these proteins are richer with euchromatin than with heterochromatin.
The acidic proteins restore the DNA-dependent
RNA synthesis, inhibited by histone, which enCopyright
© 2013 SciRes. CellBio
Z. A. KARALYAN ET AL. 129
hance the transcription of chromatin in vitro and the activation
of genes. Acidic proteins, in contrast to the histones
are tissue specific and bind to DNA only within the
tissue from which they are allocated [12,13]. Taking into
account of our data it should be concluded that the activation
of the transcription processes are in the range of 2
- 4 HPI.
Nucleolar acidic proteins―are a large group of proteins
associated with the functional activity of the nucleolar
proteins and the equivalent argentophilic proteins
[14]. The most important argentophilic proteins: RNA
polymerase I, transcription factor UBF, nucleolin (C-23),
nucleophosmin (nyumatrin or B-23), etc. In contrast to
the widespread methods of silver protein, the quantitative
cytochemical determination of acid proteins, allows us
more accurate identify variations in protein content, and
hence the functional activity of the nucleolus [15-17].
The role of these proteins in the replication of EMCV is
unquestionable, since it is next to nucleophosmin (B-23)
are localized EMCV proteins-the 2A, 3B (VPg), 3C (pro)
and 3D (pol).
It should be noted that the EMCV does not need nuclear
structures sensitive cells, as it is able to replicate in
a cell-free medium containing the individual components
of the damaged cells in the lysates [18]. Consequently,
for successful replication the virus does not require
modification of nucleolar apparatus.
We can therefore conclude that any changes in the nucleoli
are the result of the reaction of cells to viral infection
or virus modifies the activity of the nucleolus to the
successful suppression of cellular antiviral mechanisms.
In our experiments, an elevated level of the acidic
proteins in the nucleoli was preceded by the increased
content of RNA. The time gaps were approximately 1 - 2
hours. RNA synthesis in the nucleoli is directly related to
the formation of the nucleoli. It should be noted that in
the temporal boundaries the biosynthesis of the nucleoli
fits into a space equivalent to about 1 hour. 7 - 10 minutes
is the transcription of the rDNA, the synthesis of the
small subunit of the ribosome―15 - 30 min, the synthesis
of the large subunit of the ribosome―20 - 40 minutes
[19]. It is possible that identified by us the time difference
in the content of the acidic proteins and RNA in
the nucleolus is associated with the time of the biosynthesis
of ribosomes.

 发给朋友
发给朋友 分享到朋友圈
分享到朋友圈 回顶部
回顶部